
![]()
![]()
![]()
![]()
![]()
![]()
|
|
| [an error occurred while processing this directive] |
Model of cholesterol biosynthesis regulationCholesterol, an amphipathic lipid, is an essential structural component of the cell membrane and outer layer of lipoproteins of blood plasma. Simultaneously, cholesterol is a precursor of several other steroids, namely, corticosteroids, sex hormones, bile acids, and vitamin D. Cholesterol is synthesized from acetyl-CoA, and its major fraction in the blood plasma resides in the low-density lipoproteins (LDL). Cholesterol is removed from tissues with involvement of high-density lipoproteins (HDL) to be transported into the liver and transformed there into bile acids. In pathology, cholesterol is a factor causing atherosclerosis of vital cerebral arteries, heart muscle, and other organs. A high value of the LDL cholesterol to HDL cholesterol ratio in the plasma is observed during coronary atherosclerosis. This determines a great biomedical and applied importance of studying cholesterol turnover in the organism. The gene network regulating intracellular cholesterol biosynthesis is now studied in sufficient detail. Data on its performance patterns are accumulated in the GeneNet database. Acetyl-CoA is the source of all the carbon atoms of the cholesterol molecule. Cholesterol biosynthesis pathway comprises numerous stages and is controlled by a variety of enzymes, including HMG-CoA reductase, farnesyl diphosphate synthetase, and squalene synthetase. Syntheses of the enzymes listed are activated by SREBP (sterol regulatory element-binding protein). In turn, the activity of SREBP depends on the intracellular cholesterol concentration in a negative feedback mode: the lower is the concentration of metabolically active cholesterol in the cell, the higher is the SREBP activity. We have developed a model of functional dynamics of this gene network. The model describes all the stages of cholesterol biosynthesis shown in GeneNet database. In addition, the model describes the mechanisms underlying the interchange of intracellular and blood cholesterols. Negative feedbacks whereby cholesterol exerts the control over its own synthesis and synthesis of LDL receptors at the transcription level [Wang et al., 1994] are also considered. Totally, the model comprises 65 elementary processes. The model of performance dynamics of this gene network described in the GeneNet database contains 40 products (dynamic variables). Brief descriptions of the gene network objects presented in the model as dynamic variables given in the table. Table. The objects of gene network of cholesterol biosynthesis regulation presented in the models as dynamic variable
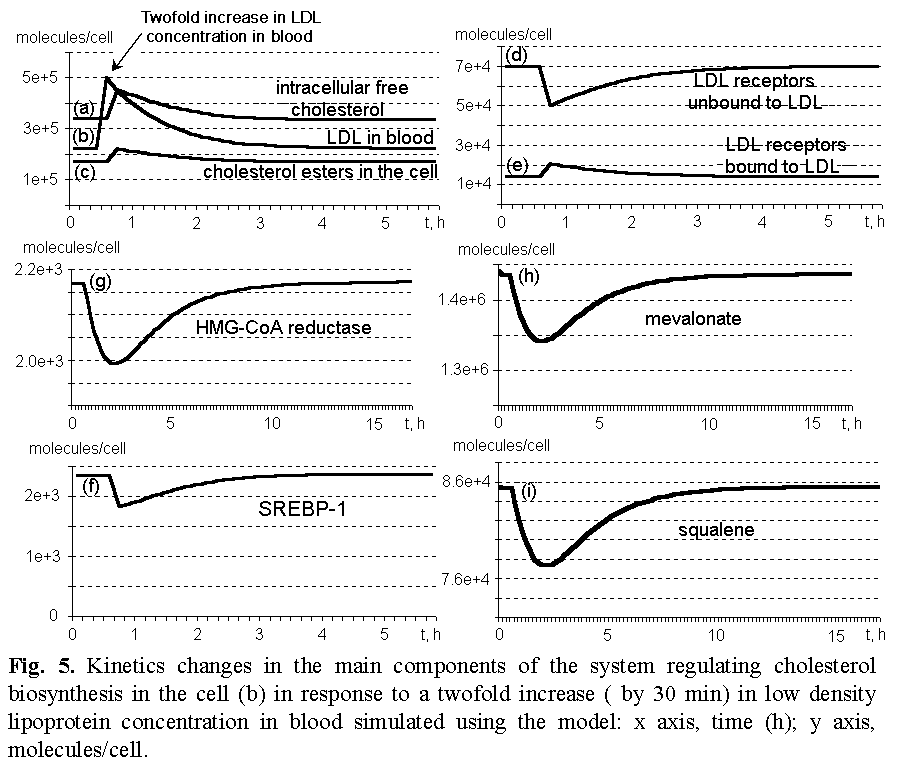
The model contains 93 constants. Values of a number of constants were assessed using the relevant published data [Gil et al., 1981; Kleinsek et al, 1979; Reed et al., 1975; Rokosz et al., 1994; Huth et al., 1975; Sasiak and Rilling, 1988; Croteau and Purkett, 1989; Barnard and Popjak, 1981; Balliano et al., 1992; Chang et al., 1998; Lundeen and Savage, 1990]. The rest parameters were determined through numerical experiments [Ratushny et al., 2000à] using quantitative and qualitative characteristics known the literature as criteria of their adequacies. This model allows the equilibrium state of the biosystem to be calculated. The equilibrium persists while the environmental conditions remain constant. If they change, for example, the content of LDL particles in blood plasma increases twofold, the system equilibrium is disturbed (Fig. 5). Consequently, the concentration of receptors bound to LDL increases (e); unbound, decreases (d). Intracellular concentrations of free cholesterol (a) and its esters (c) increase, etc. Unless a new intervention occurs, the negative feedbacks restore the initial state of the system: the initial cholesterol concentration in the cell reestablishes in approximately 3h, while the overall initial state of the system restores in 10- 15 h. BibliographyGil G., Sitges M., Hegardt, F. G. Purification and
Properties of Rat Liver Hydrohymethylglutaryl Coenzyme A Reductase phosphatases. BIOCHIMICA
ET BIOPHYSICA ACTA, 1981, 663(1), 211-221. Kleinsek DA, Porter JW An alternate method of
purification and properties of rat liver -hydroxy-methylglutaryl coenzyme A reductase. Journal
of Biological Chemistry, 1979, 254(16), 7591-7599. Reed W. D., Clinkenbeard K. D., Lane M. D. Molecular
and catalytic properties of mitochondrial (ketogenic) 3-hydroxy-3-methylglutaryl coenzyme
A synthase of liver. Journal of Biological Chemistry, 1975, 250(8), 3117-3123. Rokosz L. L., Boulton D. A., Butkiewicz E. A.,
Sanyal G., Cueto M. A., Lachance P. A., Hermes J. D. Human cytoplasmic
3-hydroxy-3-methylglutaryl Coenzyme A synthase: expression, purification, and
characterization of recombinant wild-type and Cys129 mutant enzymes. Archives of
Biochemistry and Biophysics, 1994, 312(1), 1-13. Huth W., Jonas R., Wunderlich I., Seubert W. On the
mechanism of ketogenesis and its control. Purification, kinetic mechanism and regulation
of different forms of mitochondrial acetoacetyl-CoA thiolases from ox liver. European
Journal of Biochemistry, 1975, 59(2), 475-489. Sasiak K., Rilling H. C. Purification to
homogeneity and some properties of squalene synthetase. Archives of Biochemistry and
Biophysics, 1988, 260(2), 622-627. Croteau R., Purkett P. T. Geranyl pyrophosphate
synthase: Characterization of the enzyme and evidence that this chain-length specific
prenyltransferase is associated with monoterpene biosynthesis in sage (Salvia
officinalis). Archives of Biochemistry and Biophysics, 1989, 271(2), 524-535. Barnard G. F., Popjak G. Human liver
prenyltransferase and its characterization. Biochimica et Biophysica Acta, 1981,
661(1), 87-99. Balliano G., Viola F., Ceruti M., Cattel L.
Characterization and partial purification of squalene-2,3-oxide cyclase from Saccharomyces
cerevisiae Arch.Biochem.Biophys., 1992, 293(1), 122-129. Chang C. C. Y., Lee C.-Y. G., Chang E. T., Cruz J.
C., Levesque M. C., Chang T.-Y. Recombinant acyl-CoA:cholesterol acyltransferase-1
(ACAT-1) purified to essential homogeneity utilizes cholesterol in mixed micelles or in
vesicles in a highly cooperative manner. Journal of Biological Chemistry, 1998,
273(52), 35132-35141. Lundeen S.G., Savage D.C. Characterization and
purification of bile salt hydrolase from Lactobacillus sp. strain 100-100. Journal of
Bacteriology, 1990, 172(8), 4171-4177. Ratushny A.V., Ignatieva E.V.,
Matushkin Yu.G., Likhoshvai V.A. Mathematical model of cholesterol biosynthesis regulation
in the cell. Proceeding of the second international conference on bioinformatics or genome
regulation and structure, Novosibirsk, 2000a, V.1, 199-202. Wang X., Seto R., Brown M.S. et al. SREBP-1, a membrane-bound
transcription factor released by sterol regulated proteolysis. Cell, 1994, V. 77,
53-62. |
{kind=link}