| |
|
| HOME | SITE MAP | GENENET DATABASE | PLANT-TRRD | PLANT ONTOLOGY |
| Photomorphogenesis Gene Network | Nodulation Gene Network | Flower development Gene Network |
Flower development Gene NetworkThe Gene Network of the Flower Development is available via the Internet Nadejda A. Omelyanchuk e-mail: nadya@bionet.nsc.ru See Images of gene network components Considerable volume of data has been so far accumulated on expression regulation of the genes involved in the gene network of Arabidopsis flower development. The computer system GeneNet provides convenient tools for accumulation, systematization, regular updating, and graphical representation of these data. GeneNet is applicable to processes in which regulation of gene expression has been demonstrated by experimental data on transcription, binding of transcription factors is shown in details as well as the resulting changes in gene expression and the following processes. Unfortunately, these levels of regulation of the flower development are not yet clearly understood. Nevertheless, other experimental data giving evidences on regulation of these processes in an indirect manner, such as changes in mRNA expression in mutant and transgenic lines and resulting changes in the development allowed a virtual model of this gene network, although presumably not so evident and comprehensive, but still informative, to be created. All the data involved in constructing this gene network were extracted from publications. The database contains a statement that "a gene A regulates directly the expression of a gene B" only if it is known that the transcription factor A binds to the regulatory site housed in the promoter or intron of the gene B and this results in a change in the expression of gene B. It is stated that "a gene A regulates indirectly the expression of a gene B" only if the data on at least two types of the experiments listed below are available:
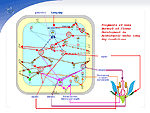
The genes lacking experimental evidences on the regulatory relations listed above were not included into the database. In the case of redundant actions of genes and co-regulators combined with an unknown mechanism of interactions between these genes, the effect of the gene set is depicted in the gene network as the effect of the major gene whose mutation changes expression of the gene regulated. The references used for gene network construction are omitted, as they all are listed with the GeneNet database. When an increase in the daylight induces flowering, the light signal via photoreceptors is transmitted to the system of light signal transduction and to the biological clock system. The last system includes the genes GIGANTEA (GI) and LATE ELONGATED HYPOCOTYL (LHY) , whose activities are altered under long day conditions, resulting in a change in the activity of gene CONSTANS (CO) . The last gene is a "middleman" between the biological clock system and gene network of floral development. Expression of gene CO in the vegetative meristem initiates and maintains the expressions of two genes-TERMINAL FLOWER1 (TFL1) and LEAFY (LFY). TFL1 is the major gene detaining the meristem at the vegetative stage through complete inhibition of the gene AP1 activity and restraining LFY to a particular activity level. A long day-stimulated increase in the gene CO activity results in considerable elevation in the activities of two genes-FT (FLOWERING LOCUS T) and LFY-in the cells of peripheral vegetative meristem. FT induces the activity of gene AP1 in an indirect manner. LFY encodes a transcription factor and is the major gene in the floral development process. Rapid initial activation of the floral development gene network and transition of flank meristem cells to formation of flower primordia requires considerable amount of the protein LFY. This is achieved due to a positive feedback circuit of the genes LFY and APETALA1 (AP1). High levels of LFY and APETALA1 proteins also underlie the irreversibility of transition to flowering, as they inhibit the gene TFL1, which suppresses the activities of these genes in vegetative meristem. This positive feedback regulatory circuit maintains the gene LFY, the major gene of the floral development process, in the state of being "constantly switched on", triggering the cascade of transcription factor genes AP1, AP3, PI, and AG. These transcription factors provide formation of various flower primodria, namely sepals (AP1), petals (AP1, AP3, and PI), stamens (AP3, PI, and AG), and carpel (AG). The major regulator of other significant positive feedback regulatory circuit supplemented with autoregulation is heterodimeric protein AP3/PI, involved in maintaining the transcription of both AP3 and PI genes, coding for its monomers. It has been demonstrated that this heterodimer is a component of the tetramer ŔĐ3/PI/SEP3/AP1, which increases the gene AP3 transcription and is the major factor providing formation of the petal primodria. As for stamen primodria, the key factor here is the tetramer ŔĐ3/PI/SEP3/AG. Heterodimer AP3/PI also participate in switching on the genes SUPERMAN (SUP), responsible for forming the border between stamens and carpels, and NAP, providing the further development of stamens and petals. A graphical layout of the gene network in question makes the relation between the time course of transcription regulation and developmental processes in question more illustrative. The gene network of flower development in Arabidopsis has a number of characteristics common for gene networks of developmental processes, such as positive feedback circuits and cassette-type gene activation. Our computer system presents the current experimental data on regulation of gene expression in flower development and provides their updating with new information published on already described and novel genes on a regular basis. |
||
| ©1997-2002, IC&G SB RAS, Laboratory of Theoretical Genetics |
Web-master Ekaterina Denisova |